Recombinant Human Transcriptional repressor CTCF (CTCF), partial
-
中文名稱:Recombinant Human Transcriptional repressor CTCF(CTCF),partial
-
貨號(hào):CSB-YP006138HU1
-
規(guī)格:
-
來(lái)源:Yeast
-
其他:
-
中文名稱:Recombinant Human Transcriptional repressor CTCF(CTCF),partial
-
貨號(hào):CSB-EP006138HU1
-
規(guī)格:
-
來(lái)源:E.coli
-
其他:
-
中文名稱:Recombinant Human Transcriptional repressor CTCF(CTCF),partial
-
貨號(hào):CSB-EP006138HU1-B
-
規(guī)格:
-
來(lái)源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Human Transcriptional repressor CTCF(CTCF),partial
-
貨號(hào):CSB-BP006138HU1
-
規(guī)格:
-
來(lái)源:Baculovirus
-
其他:
-
中文名稱:Recombinant Human Transcriptional repressor CTCF(CTCF),partial
-
貨號(hào):CSB-MP006138HU1
-
規(guī)格:
-
來(lái)源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:Greater than 90% as determined by SDS-PAGE.
-
基因名:
-
Uniprot No.:
-
別名:CTCF; Transcriptional repressor CTCF; 11-zinc finger protein; CCCTC-binding factor; CTCFL paralog
-
種屬:Homo sapiens (Human)
-
蛋白長(zhǎng)度:Partial
-
表達(dá)區(qū)域:266-727aa
-
氨基酸序列FQCELCSYTCPRRSNLDRHMKSHTDERPHKCHLCGRAFRTVTLLRNHLNTHTGTRPHKCPDCDMAFVTSGELVRHRRYKHTHEKPFKCSMCDYASVEVSKLKRHIRSHTGERPFQCSLCSYASRDTYKLKRHMRTHSGEKPYECYICHARFTQSGTMKMHILQKHTENVAKFHCPHCDTVIARKSDLGVHLRKQHSYIEQGKKCRYCDAVFHERYALIQHQKSHKNEKRFKCDQCDYACRQERHMIMHKRTHTGEKPYACSHCDKTFRQKQLLDMHFKRYHDPNFVPAAFVCSKCGKTFTRRNTMARHADNCAGPDGVEGENGGETKKSKRGRKRKMRSKKEDSSDSENAEPDLDDNEDEEEPAVEIEPEPEPQPVTPAPPPAKKRRGRPPGRTNQPKQNQPTAIIQVEDQNTGAIENIIVEVKKEPDAEPAEGEEEEAQPAATDAPNGDLTPEMILSMMDR
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Chromatin binding factor that binds to DNA sequence specific sites. Involved in transcriptional regulation by binding to chromatin insulators and preventing interaction between promoter and nearby enhancers and silencers. Acts as transcriptional repressor binding to promoters of vertebrate MYC gene and BAG1 gene. Also binds to the PLK and PIM1 promoters. Acts as a transcriptional activator of APP. Regulates APOA1/C3/A4/A5 gene cluster and controls MHC class II gene expression. Plays an essential role in oocyte and preimplantation embryo development by activating or repressing transcription. Seems to act as tumor suppressor. Plays a critical role in the epigenetic regulation. Participates in the allele-specific gene expression at the imprinted IGF2/H19 gene locus. On the maternal allele, binding within the H19 imprinting control region (ICR) mediates maternally inherited higher-order chromatin conformation to restrict enhancer access to IGF2. Plays a critical role in gene silencing over considerable distances in the genome. Preferentially interacts with unmethylated DNA, preventing spreading of CpG methylation and maintaining methylation-free zones. Inversely, binding to target sites is prevented by CpG methylation. Plays an important role in chromatin remodeling. Can dimerize when it is bound to different DNA sequences, mediating long-range chromatin looping. Mediates interchromosomal association between IGF2/H19 and WSB1/NF1 and may direct distant DNA segments to a common transcription factory. Causes local loss of histone acetylation and gain of histone methylation in the beta-globin locus, without affecting transcription. When bound to chromatin, it provides an anchor point for nucleosomes positioning. Seems to be essential for homologous X-chromosome pairing. May participate with Tsix in establishing a regulatable epigenetic switch for X chromosome inactivation. May play a role in preventing the propagation of stable methylation at the escape genes from X- inactivation. Involved in sister chromatid cohesion. Associates with both centromeres and chromosomal arms during metaphase and required for cohesin localization to CTCF sites. Regulates asynchronous replication of IGF2/H19. Plays a role in the recruitment of CENPE to the pericentromeric/centromeric regions of the chromosome during mitosis.
-
基因功能參考文獻(xiàn):
- The authors propose that cellular CCCTC-binding factor binding at the herpes simplex virus 1 CCCTC-binding factor binding sites (CTRL2) acts as a chromatin insulator to keep viral chromatin in a form that is poised for reactivation, a state which we call poised latency. PMID: 29437926
- neither the deletion of the CTCF locus nor the ectopic insertion of Firre cDNA or its ectopic expression are sufficient to alter topologically associated domains in a sex-specific or allele-specific manner PMID: 29654311
- CTCF maintains regulatory homeostasis of cancer pathways. PMID: 30086769
- these results suggest that CTCF participate in DNA damage response via poly(ADP-ribosylation). PMID: 28262757
- Studies suggest that the connection between DNA-binding protein CTCF (CTCF), cohesin, chromatin structure, and behavior is important in understanding of the development of behavior in general, and neurodevelopmental disorders in particular [Review]. PMID: 29110030
- ID1, CTCF and ELK1 may be associated with prostate cancer, and may be potential therapeutic targets for the treatment of this disease. PMID: 29956775
- CTCF promotes HSV-1 lytic transcription by facilitating the elongation of RNA Pol II and preventing silenced chromatin on the viral genome. PMID: 28045091
- These results, together with a prior exomesequencing based study, suggest that CTCF mutations may be involved in the development of ovarian endometriosis. PMID: 29845264
- findings establish for the first time that CTCF is an important regulator of the homologous recombination repair pathway. PMID: 28560323
- Findings indicate that CCCTC-binding factor (CTCF)-driven doublesex and mab-3 related transcription factor 2 protein (TERRA) transcription acts in cis to facilitate telomere repeat replication and chromosome stability. PMID: 29235471
- poly(ADP-ribosyl)ated CTCF changes its DNA binding and localisation in a breast cell line which is associated with nucleosome repositioning. PMID: 29981477
- Here, we show that PARP1 and host insulator protein CTCF colocalize at specific sites throughout the EBV genome and provide evidence to suggest that PARP1 acts to stabilize CTCF binding and maintain the open chromatin landscape at the active Cp promoter during type III latency. Further, PARP1 activity is important in maintaining latency type-specific viral gene expression. PMID: 29976663
- HOTTIP cooperates with CTCF to coordinate HOXA gene expression. PMID: 29698677
- CD4(+) T cells showed the greatest increase (threefold) in ORMDL3 expression in individuals carrying the asthma-risk alleles, where ORMDL3 negatively regulated interleukin-2 production. The asthma-risk variants rs4065275 and rs12936231 switched CTCF-binding sites in the 17q21 locus. PMID: 27848966
- Our data reveal that vigilin is essential for maintenance of imprinting of IGF2 gene via functional interaction between KH1-7 domains of vigilin and zinc-finger domains of CTCF. PMID: 29157910
- This study confirms that haploinsufficiency of CTCF causes distinct clinical features, and that a microdeletion encompassing CTCF could cause a recognisable CTCF deletion syndrome. Perturbed DNA methylation at CTCF binding sites, not at imprinted loci, may underlie the pathomechanism of the syndrome. PMID: 28848059
- structural studies show that the sequence-specific interactions between zinc fingers and CTCF-binding sites determine the directionality and conservation of CTCF recognition. PMID: 29076501
- CTCF may be a key factor that contributes to gene co-mutations in cancer. PMID: 27762310
- results support a model in which YY1 acts as an architectural protein to connect developmentally regulated looping interactions; the location of YY1-mediated interactions may be demarcated in development by a preexisting topological framework created by constitutive CTCF-mediated interactions. PMID: 28536180
- The MeCP2, a protein whose mutated forms are involved in Rett syndrome; and CTCF, a constitutive transcriptional insulator. PMID: 28796949
- The results show that cohesin has an essential genome-wide function in mediating long-range chromatin interactions and support the hypothesis that cohesin creates these by loop extrusion, until it is delayed by CTCF in a manner dependent on PDS5 proteins, or until it is released from DNA by WAPL. PMID: 29217591
- CTCF-FOXM1 axis regulates tumour growth and metastasis in hepatocellular carcinoma cells. PMID: 28862757
- we show CTCF binding site mutations to be functional by demonstrating allele-specific reduction of CTCF binding to mutant alleles. While topologically associating domains with mutated CTCF anchors in melanoma contain differentially expressed cancer-associated genes, CTCF motif mutations appear generally under neutral selection PMID: 27974201
- CTCF-mediated long-range interactions are integral for a multitude of topological features of interphase chromatin, such as the formation of topologically associated domains, domain insulation, enhancer blocking and even enhancer function. PMID: 26802288
- Authors have identified two novel pro-tumorigenic roles (promoting cell survival and altering cell polarity) for genetic alterations of CTCF in endometrial cancer. PMID: 28319062
- Describe several protein-DNA complex structures of a human CTCF tandem zinc-finger array, explaining the adaptability of CTCF to sequence variations and the positiondependent effect of differential DNA methylation at two cytosine residues, and revealing a potential function of C-terminal ZF8 and ZF9 spanning across the DNA phosphate backbone. PMID: 28529057
- CCCTC-binding factor (CTCF) targets the binding sites within MYCN promoter to facilitate its expression in neuroblastoma (NB) cells. PMID: 26549029
- we review recent high-resolution chromosome conformation capture and functional studies that have informed models of the spatial and regulatory compartmentalization of mammalian genomes, and discuss mechanistic models for how CTCF and cohesin control the functional architecture of mammalian chromosomes. PMID: 27089971
- GAD1 is reactivated by DNA methylation, which provided a model for DNA methylation and the active orchestration of oncogenic gene expression by CTCF in cancer cells. PMID: 26549033
- epigenetic factor CTCF-mediated chromatin remodeling regulates interactions between eye-specific PAX6 and those genes that are induced/associated with cell differentiation to modulate corneal epithelial cell-specific differentiation PMID: 27583466
- Rta-mediated decreased binding of CTCF in the viral genome is concurrent with virus reactivation. Via interfering with CTCF binding, in the host genome Rta can function as a transcriptional repressor for gene silencing. PMID: 28490592
- although we were unable to detect HD-associated DNA methylation alterations at queried sites, we found that DNA methylation may be correlated to the age of disease onset in cortex tissues. Moreover, our data suggest that DNA methylation may, in part, contribute to tissue-specific HTT transcription through differential CTCF occupancy. PMID: 26953320
- vitamin D-sensitive CTCF sites provide further mechanistic details to the epigenome-wide understanding of 1,25(OH)2D3-mediated gene regulation PMID: 27569350
- These findings indicate that erythroid specific activator GATA-1 acts at CTCF sites around the beta-globin locus to establish tissue-specific chromatin organization. PMID: 28161276
- TOP2B is positioned to solve topological problems at diverse cis-regulatory elements and its occupancy is a highly ordered and prevalent feature of CTCF/cohesin binding sites that flank TADs. PMID: 27582050
- CTCF binding to eRNAs and promoters is facilitated by estrogen when chromatin establishes contacts with nuclear lamina. PMID: 27638884
- we investigated the cell-type specificities of CTCF sites related to these functions across five cell types. Our study provides new insights into the multivalent functions of CTCF in the human genome PMID: 27067545
- The results indicate that the initial chromatin conformation affects subsequent RA-induced HOXA gene activation. Our study uncovers that a removable insulator spatiotemporally switches higher-order chromatin and multiple gene activities via cooperation of CTCF and key transcription factors. PMID: 27798106
- we find no evidence for selection driving these distinctive patterns of mutation. The mutational load at CTCF-binding sites is substantially determined by replication timing and the mutational signature of the tumor in question, suggesting that selectively neutral processes underlie the unusual mutation patterns. PMID: 27490693
- data demonstrate a reciprocal relationship between Hmgb2 and Ctcf in controlling aspects of chromatin structure and gene expression. PMID: 27226577
- Study described the formation of mutually exclusive complexes of ENY2 with insulator proteins and Sgfl1-a component of the SAGA complex, direct binding partner for ENY2 PMID: 27417714
- This is supported by the depletion of CTCF in glioblastoma cells affecting the expression levels of NOTCH2 as a target of miR-181c. CONCLUSION: Together, our results point to the epigenetic role of CTCF in the regulation of microRNAs implicated in tumorigenesis PMID: 26983574
- Our data show that aberrant epigenetic inactivation of DUSP2 occurs in carcinogenesis and that CTCF is involved in the epigenetic regulation of DUSP2 expression. PMID: 26833217
- Mutational analysis highlighted a potential role for CTCF, a crucial regulator of long-range chromatin interactions, in head and neck cancer progression. PMID: 26747525
- CTCF and cohesins shape the genome during evolution. (Review) PMID: 26439501
- The action of SNF2H at CTCF sites is functionally important as depletion of CTCF or SNF2H affects transcription of a common group of genes. PMID: 27019336
- CSB and CTCF can regulate each other's chromatin association, thereby modulating chromatin structure and coordinating gene expression in response to oxidative stress. PMID: 26578602
- CTCF binds to the provirus at a sharp border in epigenetic modifications in the pX region of the HTLV-1 provirus in T cells naturally infected with HTLV-1. This may cause widespread abnormalities in host cell chromatin structure and gene expression. PMID: 26929370
- CTCF/cohesin coordinates HOXA cluster higher-order chromatin structure and expression during development PMID: 26376810
- CTCF has a role in regulating SLC45A3-ELK4 Chimeric RNA PMID: 26938874
顯示更多
收起更多
-
相關(guān)疾病:Mental retardation, autosomal dominant 21 (MRD21)
-
亞細(xì)胞定位:Nucleus, nucleoplasm. Chromosome. Chromosome, centromere. Note=May translocate to the nucleolus upon cell differentiation. Associates with both centromeres and chromosomal arms during metaphase. Associates with the H19 ICR in mitotic chromosomes. May be preferentially excluded from heterochromatin during interphase.
-
蛋白家族:CTCF zinc-finger protein family
-
組織特異性:Ubiquitous. Absent in primary spermatocytes.
-
數(shù)據(jù)庫(kù)鏈接:
Most popular with customers
-
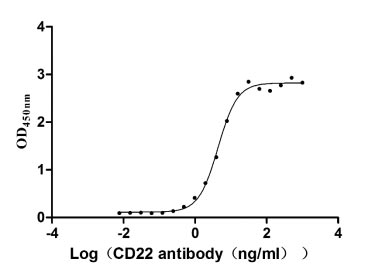
Recombinant Human B-cell receptor CD22 (CD22), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
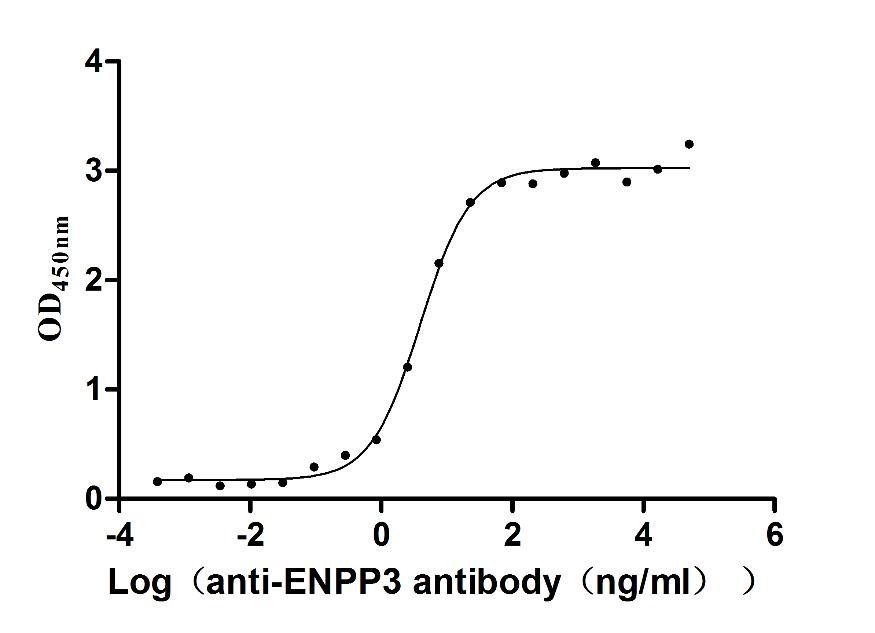
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
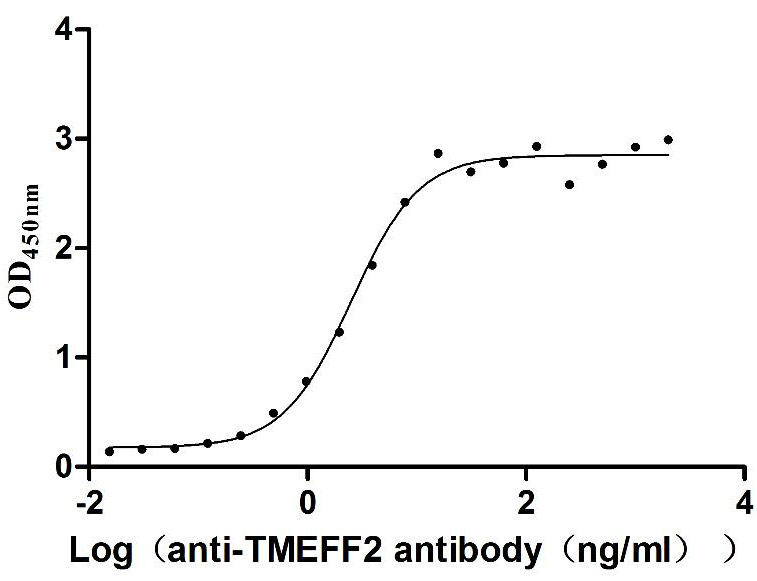
Recombinant Human Tomoregulin-2 (TMEFF2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human C-C chemokine receptor type 8 (CCR8)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Macaca fascicularis Dipeptidase 3(DPEP3) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-

Recombinant Human Mucin-13(MUC13),partial (Active)
Express system: yeast
Species: Homo sapiens (Human)











